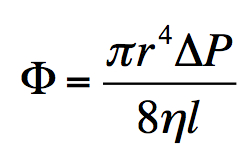This roadside sign (“Many Species Live Here”) is from the Rincon de la Vieja Rainforest Reserve*, in Guanacaste, Costa Rica, where I have been fortunate enough to do a bit of research on forest biodiversity. But it might just as well represent Earth in its entirety.
But how many species do live on Earth?
This is one of those big questions, perhaps the big question of biodiversity. And as with most big questions, it has many answers, the first of which is, quite simply, “We don’t know.”
However, this typical small answer to big questions is also terribly unsatisfying. So if we want to struggle on against uncertainty, how can we go about estimating the number of species on Earth? First, it is important to note that we can’t simply count up all of the names published by expert taxonomists. The named species total up to about 1.5 million, but while such lists are fairly complete for some groups of organisms, like mammals, for others, like fungi or nematode worms, they are woefully spotty. Scientists have simply not had the time and resources necessary to describe and name all of the organisms. In fact, even this year a new species of monkey was described from the central African rain forests, so we’re still getting to know even some of our closest relatives on Life’s family tree.
So where do we start? Sometimes in science, it pays to start with the obvious, and work from there. Back in 1988, a physicist turned ecologist named Robert May (now Lord May – perhaps the most ennobled ecologist) used a paper in the journal Nature to outline the kind of information that would be relevant to making such an estimate. He pointed out that most of the undescribed species tend to be small and/or rare, either in the sense of having low overall abundance or by being endemic to a very particular environment, because small, rare creatures are hard to find. Sounds obvious to me, but it highlights some of the factors we might want to consider.
Focusing on size, May pointed out that overall, there are generally many more small species than large species, but that the pattern shifts, declining for critters under about a centimeter long. If the decline observed among small species was only because of undersampling, May extrapolated that there could be anywhere from 10-50 million terrestrial animal species alone. While May admits the questionable nature of this estimate, the point remains that there may be many, many animal species out there waiting to be described, especially among the smallest critters. Indeed, several recently discovered vertebrates have been record-setting miniatures, including a fly-sized frog from New Guinea that could easily sit on your pinkie nail and four species of similarly sized chameleons from Madagascar.
Another option is to go to a place where many undescribed species are likely to dwell, sample them extensively, then extrapolate to other unsampled regions. This approach was pioneered by the entomologist Terry Erwin, who in the late 1970’s, went down to Panama to study rain forest beetles. (You’re probably starting to see the pattern that most of the undescribed terrestrial species are from the tropics; the reasons for that will be the subject of another post.) Since most of the beetles in a rainforest live high in the canopy, Erwin had to bring them down to the ground for study, so he constructed large tents around 19 trees belonging to a single, relatively common tree species, Luehea seemannii, then fogged the enclosed trees with an insecticide. Combing through the insects that rained down from the canopy, Erwin found about 1,200 different beetle species. But Erwin didn’t just want to know how many beetle species were on one tree species; he wanted to know how many insect species there were in the world’s tropical forests.
So once he’d killed them all and sorted them out, he did some extrapolating. First, he assumed that beetles were about 2/5 of all arthropods, the group that includes insects, spiders, etc. This is a pretty well-supported number; nature does seem to love beetles. Having a fair amount of experience hunting for tropical beetles, he also estimated that about 2/3 of them lived in the canopy, while the rest lived low on the trunk (or inside it), inside the leaves, or down in the soil among the roots. Being an expert on beetles, he was able to estimate that about 13.5% of the species he found were specialists, living only in the canopy of Luehea seemannii. Putting these numbers together, and assuming that other insect groups were similar in the diversity of their habitats and their degree of specialization, he estimated that their were about 611 specialist insects on a single tropical rainforest tree species. Taking the next, more daring step, given that there are an estimated 50,000 tree species in Earth’s tropical rain forests, Erwin ended up with a global estimate of about 30 million tropical insects.
Now given all of the attendant assumptions, Erwin’s number was not meant to be a hard-and-fast estimate. Nor, for that matter, was May’s. Instead, they are meant to put some bounds on our thinking, based on a set of rational, and of course debatable, assumptions. Compiling these sorts of estimates for different types of organisms and consulting taxonomic experts for their opinions has lead to a best guess of there being between 5 and 300 million species on Earth. The very broadness of this answer makes it almost as unsatisfying to me as “we don’t know.” In any case, it points to the fact that we still have a wealth of biodiversity for taxonomists to discover. Moreover, we could really use some better methods for narrowing down our estimates.
A recent (2011) paper by Camilo Mora and colleagues attempts to provide such a narrower estimate. Instead of just considering the number of species, Mora’s group took an historical approach and looked at the rates at which new species were described. This angle had been taken on biodiversity estimation before, but they expanded the approach by capitalizing on the fact that while scientists have certainly not described all of Earth’s species, we have a pretty good handle on the number of some “higher taxa,” meaning higher in the taxonomic hierarchy you probably learned in school: kingdom, phylum, class, order, family, genus, species. They examined data on the rates at which new groups were described for each taxonomic level, from 1750 to today. Early in the development of biodiversity classification, new groups were accumulating quickly in the scientific record, but eventually, the numbers of most of the higher taxonomic groups, like phyla and classes, level off. Using these estimates for the number of higher taxonomic groups in a mathematical model, they are able to estimate the number of species. Across all eukaryotes (anything that is not a bacterium, archaean, or virus), they estimate that the Earth harbors between 7.4 and 10 million species, at the lower end of the 5-300 million range, but still a pretty big number. In fact, if they are right, despite 250 years of intensive study, scientists have described less than 15% of the species that call Earth home.
Like May’s and Erwin’s numbers, Mora’s is not a definitive statement, but a step along the way to further scientific insight. In the grand and messy tradition of science, their approach has attracted both support and criticism, with taxonomic, evolutionary, and statistical experts weighing in. In any case, the tall order of describing and quantifying Earth’s existing biodiversity is given added urgency by the current extinction crisis, and scientists are engaged in developing a variety of field, laboratory, and computational methods to better answer the question. So while we don’t actually know how many species there are on Earth, by any estimate, it’s clear that we have a lot of work to do, and a lot of it needs to be done in the tropics.
*The Rincon Reserve is part of the larger Area Conservacion Guanacaste (ACG), which is a UNESCO World Heritage Site. The ACG encompasses about 2% of Costa Rica’s land area, including tropical dry forest, rain forest, and cloud forest, as well as a substantial marine reserve. It is managed by the government of Costa Rica for conservation and scientific research. It houses about 60% of the species that occur in Costa Rica and is one of the most well-surveyed, and beautiful tropical regions on Earth. Find out more about the ACG (in Spanish) or learn why you should donate to the Guanacaste Dry Forest Conservation Fund (which also protects rain forest and cloud forest (in English).